徐州廣州細(xì)胞外基質(zhì)膠
- 產(chǎn)地
- 蘇州
- 品牌
- 細(xì)胞外基質(zhì)膠
- 型號(hào)
- 齊全
- 是否定制
- 是
此現(xiàn)象稱為凋亡.不同的細(xì)胞外基質(zhì)對(duì)細(xì)胞增殖的影響不同.例如,成纖維細(xì)胞在纖粘連蛋白基質(zhì)上增殖加快,在層粘連蛋白基質(zhì)上增殖減慢;而上皮細(xì)胞對(duì)纖粘連蛋白及層粘連蛋白的增殖反應(yīng)則相反.部位細(xì)胞的增殖喪失了定著依賴性,可在半懸浮狀態(tài)增殖。如何理解細(xì)胞外基質(zhì)影響細(xì)胞的粘附過程:影響細(xì)胞的存活、生長(zhǎng)與死亡正常真核細(xì)胞,除成熟血細(xì)胞外,大多須粘附于特定的細(xì)胞外基質(zhì)上才能克制凋亡而存活,稱為定著依賴性.例如,上皮細(xì)胞及內(nèi)皮細(xì)胞一旦脫離了細(xì)胞外基質(zhì)則會(huì)發(fā)生程序性死亡.什么是細(xì)胞外基質(zhì):多細(xì)胞有機(jī)體中,細(xì)胞周圍由多種大分子組成的復(fù)雜網(wǎng)絡(luò),稱作細(xì)胞外基質(zhì)。徐州廣州細(xì)胞外基質(zhì)膠

細(xì)胞外基質(zhì)生理學(xué)功能:剛度和彈性:從柔軟的腦組織到堅(jiān)硬的骨組織,細(xì)胞外基質(zhì)可以以不同程度的剛度和彈性存在。細(xì)胞外基質(zhì)的彈性可以相差幾個(gè)數(shù)量級(jí)。這一特性主要取決于膠原蛋白和彈性蛋白的濃度,較近已證明細(xì)胞外基質(zhì)的剛度和彈性這個(gè)特性在調(diào)節(jié)許多細(xì)胞功能中發(fā)揮著重要作用。細(xì)胞可以通過施力和測(cè)量產(chǎn)生的反作用力來感知環(huán)境的機(jī)械特性。這發(fā)揮著重要作用,因?yàn)樗兄谡{(diào)節(jié)許多重要的細(xì)胞過程,包括細(xì)胞收縮,細(xì)胞遷移,細(xì)胞增殖,分化[和細(xì)胞死亡(凋亡)。克制非肌肉肌球蛋白ⅱ阻斷了大部分這些效應(yīng),表明它們確實(shí)與細(xì)胞外基質(zhì)的機(jī)械特性有關(guān),這在過去十年中已成為研究的新焦點(diǎn)。合肥太原細(xì)胞外基質(zhì)膠細(xì)胞外基質(zhì)主要由5類物質(zhì)組成,即膠原蛋白、非膠原蛋白、彈性蛋白、蛋白聚糖與氨基聚糖。

細(xì)胞外基質(zhì)和系統(tǒng)之間的這種相互作用在再生物種中是如何工作的尚不清楚。刺胞動(dòng)物系統(tǒng)的主要調(diào)節(jié)因子是蛋白酶、絲氨酸蛋白酶克制劑、克菌蛋白和補(bǔ)體系統(tǒng)。的原始機(jī)制是克菌肽(AMPs),在水螅體再生過程中,一些被歸類為AMPs的基因被上調(diào)。細(xì)胞外基質(zhì)不光靜態(tài)的發(fā)揮支持、連接、保水、保護(hù)等物理作用。系統(tǒng)和細(xì)胞外基質(zhì)之間的串?dāng)_:ECM是三維網(wǎng)狀,支持細(xì)胞,調(diào)節(jié)重要的細(xì)胞過程:增殖,粘附,遷移,細(xì)胞分化和炎癥。在對(duì)損傷的反應(yīng)中,**發(fā)生的事件包括系統(tǒng)的啟動(dòng)和基質(zhì)金屬蛋白酶(MMPs)的上調(diào)。細(xì)胞對(duì)損傷信號(hào)的反應(yīng)進(jìn)程和較終結(jié)果在一定程度上受創(chuàng)床中存在的特定MMP及其活性持續(xù)時(shí)間的控制。克制巨噬細(xì)胞募集到損傷部位已被證明可以克制再生;然而,在體內(nèi)對(duì)ECM重塑的影響研究較少。
細(xì)胞外基質(zhì)的組分及組裝形式由所產(chǎn)生的細(xì)胞決定,并與組織的特殊功能需要相適應(yīng)。例如,角膜的細(xì)胞外基質(zhì)為透明柔軟的片層,肌腱的則堅(jiān)韌如繩索。細(xì)胞外基質(zhì)不光靜態(tài)的發(fā)揮支持、連接、保水、保護(hù)等物理作用,而且動(dòng)態(tài)的對(duì)細(xì)胞產(chǎn)生很全影響。細(xì)胞外基質(zhì)的成分:構(gòu)成細(xì)胞外基質(zhì)的大分子種類繁多,可大致歸納為四大類:膠原、非膠原糖蛋白、氨基聚糖與蛋白聚糖、以及彈性蛋白。上皮組織、肌組織及腦與脊髓中的ECM含量較少,而結(jié)締組織中ECM含量較高。細(xì)胞和基底之間的耦合(通過粘附分子)屬于高度非線性耦合。

蛋白聚糖糖胺聚糖(GAGs)是碳水化合物聚合物,主要附著在細(xì)胞外基質(zhì)蛋白上形成蛋白聚糖(透明質(zhì)酸是一個(gè)明顯的例外,見下文)。蛋白聚糖具有吸引帶正電荷的鈉離子(Na+)的凈負(fù)電荷,鈉離子通過滲透作用吸引水分子,保持細(xì)胞外基質(zhì)和固有細(xì)胞水分充足。蛋白聚糖也有助于在細(xì)胞外基質(zhì)中捕獲和儲(chǔ)存生長(zhǎng)因子。結(jié)構(gòu):細(xì)胞外基質(zhì)的成分由固有細(xì)胞在細(xì)胞內(nèi)產(chǎn)生,并通過胞吐作用分泌到細(xì)胞外基質(zhì)中。一旦被分泌到胞外,它們就會(huì)與現(xiàn)有基質(zhì)聚集在一起。細(xì)胞外基質(zhì)由纖維蛋白和糖胺聚糖(GAGs)交聯(lián)形成的網(wǎng)狀物組成。細(xì)胞外基質(zhì)與干細(xì)胞一起生長(zhǎng)并再生人體的所有部分。貴陽細(xì)胞外基質(zhì)膠直銷價(jià)
導(dǎo)致腎小球內(nèi)腎臟細(xì)胞外基質(zhì)合成異常增加。徐州廣州細(xì)胞外基質(zhì)膠
細(xì)胞外基質(zhì):接著研究者為了實(shí)現(xiàn)不同功能EVs的隨需釋放,設(shè)計(jì)了一種由EVs組裝而成的雙層AH,將EVs與AH結(jié)合形成具有骨誘導(dǎo)的多相控釋效應(yīng)的BECM,并研究了BECM在體外對(duì)BMSCs的作用。活/死細(xì)胞染色顯示活細(xì)胞比例無明顯差異,說明BECM具有良好的生物相容性(圖4A,B)。Transwell和CCK8實(shí)驗(yàn)表明,AH+C-EVs和BECM均促進(jìn)了BMSCs的增殖和遷移。骨誘導(dǎo)7天后ALP染色及ALP活性檢測(cè)顯示,AH、AH+C-EVs、BECM均促進(jìn)了ALP的表達(dá)(圖4C-I)。為了在基因水平上評(píng)價(jià)各組成骨特性,檢測(cè)了成骨標(biāo)志物ALP、COL1A1、RUNX2在BMSCs中的表達(dá),結(jié)果顯示AH、AH+C-EVs和BECM均可促進(jìn)成骨基因。徐州廣州細(xì)胞外基質(zhì)膠
細(xì)胞外基質(zhì)和系統(tǒng)之間的這種相互作用在再生物種中是如何工作的尚不清楚。刺胞動(dòng)物系統(tǒng)的主要調(diào)節(jié)因子是蛋白酶、絲氨酸蛋白酶克制劑、克菌蛋白和補(bǔ)體系統(tǒng)。的原始機(jī)制是克菌肽(AMPs),在水螅體再生過程中,一些被歸類為AMPs的基因被上調(diào)。細(xì)胞外基質(zhì)不光靜態(tài)的發(fā)揮支持、連接、保水、保護(hù)等物理作用。系統(tǒng)和細(xì)胞外基質(zhì)之間的串?dāng)_:ECM是三維網(wǎng)狀,支持細(xì)胞,調(diào)節(jié)重要的細(xì)胞過程:增殖,粘附,遷移,細(xì)胞分化和炎癥。在對(duì)損傷的反應(yīng)中,**發(fā)生的事件包括系統(tǒng)的啟動(dòng)和基質(zhì)金屬蛋白酶(MMPs)的上調(diào)。細(xì)胞對(duì)損傷信號(hào)的反應(yīng)進(jìn)程和較終結(jié)果在一定程度上受創(chuàng)床中存在的特定MMP及其活性持續(xù)時(shí)間的控制。克制巨噬細(xì)胞募集到損傷部位...
- 蘇州正規(guī)細(xì)胞外基質(zhì)膠廠家直銷 2025-07-15
- 寧波成都細(xì)胞外基質(zhì)膠 2025-07-15
- 南昌正規(guī)細(xì)胞外基質(zhì)膠廠家推薦 2025-07-15
- 開封細(xì)胞外基質(zhì)膠價(jià)格 2025-07-15
- 唐山細(xì)胞外基質(zhì)膠單價(jià) 2025-07-15
- 溫州唐山細(xì)胞外基質(zhì)膠 2025-07-15
- 石家莊正規(guī)細(xì)胞外基質(zhì)膠哪家便宜 2025-07-14
- 深圳正規(guī)細(xì)胞外基質(zhì)膠廠家直銷 2025-07-14
- 廣州細(xì)胞外基質(zhì)膠直銷價(jià) 2025-07-14
- 杭州細(xì)胞外基質(zhì)膠廠家 2025-07-14


- 南京鄭州細(xì)胞外基質(zhì)膠 2025-07-13
- 唐山細(xì)胞外基質(zhì)膠廠家現(xiàn)貨 2025-07-13
- 北京細(xì)胞外基質(zhì)膠銷售廠家 2025-07-13
- 南京正規(guī)細(xì)胞外基質(zhì)膠直銷價(jià) 2025-07-13
- 南京正規(guī)細(xì)胞外基質(zhì)膠廠家批發(fā)價(jià) 2025-07-13
- 蘇州細(xì)胞外基質(zhì)膠廠家直銷 2025-07-13

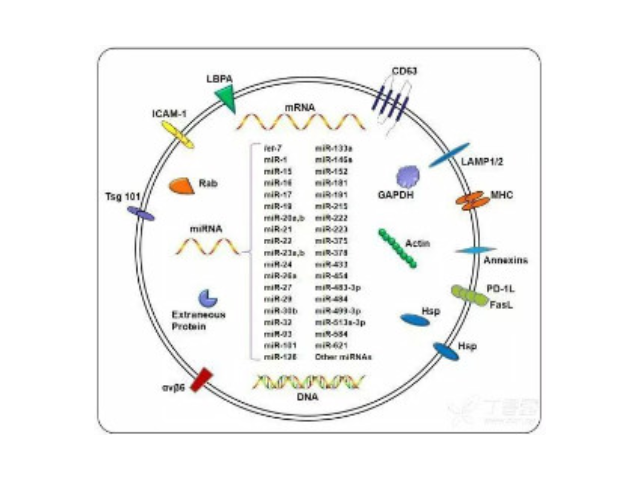

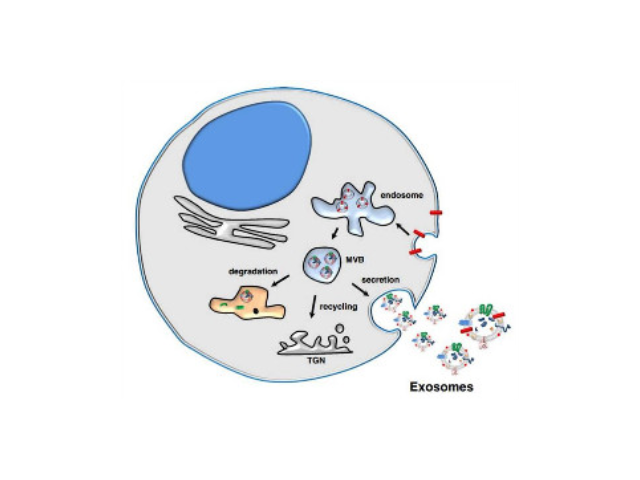

- 蘇州正規(guī)細(xì)胞外基質(zhì)膠廠家直銷 07-15
- 寧波成都細(xì)胞外基質(zhì)膠 07-15
- 南昌正規(guī)細(xì)胞外基質(zhì)膠廠家推薦 07-15
- 武漢正規(guī)外泌體提取試劑廠家 07-15
- 開封細(xì)胞外基質(zhì)膠價(jià)格 07-15
- 南京正規(guī)無血清細(xì)胞凍存液推薦廠家 07-15
- 鄭州正規(guī)外泌體提取試劑哪家便宜 07-15
- 貴陽外泌體提取試劑哪里買 07-15
- 長(zhǎng)沙無血清細(xì)胞凍存液廠家推薦 07-15
- 唐山細(xì)胞外基質(zhì)膠單價(jià) 07-15