太原正規(guī)細(xì)胞外基質(zhì)膠價(jià)格
- 產(chǎn)地
- 蘇州
- 品牌
- 細(xì)胞外基質(zhì)膠
- 型號(hào)
- 齊全
- 是否定制
- 是
這種轉(zhuǎn)化細(xì)胞接種入正常機(jī)體,常能長(zhǎng)成塊,并侵潤(rùn)正常組織,發(fā)生普遍轉(zhuǎn)移。又如上皮細(xì)胞分泌膠原蛋白和膜粘蛋白于上皮組織的基底層上,反之,這些蛋白又作為信號(hào)“指揮”上皮細(xì)胞生長(zhǎng)、遷移的方向。在胚胎發(fā)育或愈傷再生時(shí),上皮細(xì)胞正是沿著基底層發(fā)展的。由此可知,調(diào)節(jié)細(xì)胞生長(zhǎng)、發(fā)育的若干信息正是通過(guò)胞外基質(zhì)傳遞的。細(xì)胞外基質(zhì)的主要類(lèi)型及功能:對(duì)人類(lèi)細(xì)胞的研究表明,細(xì)胞外基質(zhì)中的纖粘蛋白主要由成纖維細(xì)胞、上皮細(xì)胞等分泌并附著在細(xì)胞表面,其作用是促進(jìn)細(xì)胞對(duì)基質(zhì)的貼附,細(xì)胞之間的粘著,細(xì)胞內(nèi)微絲及應(yīng)力纖維的構(gòu)建。觀察到轉(zhuǎn)化的體外培養(yǎng)的成纖維細(xì)胞,表面纖維蛋白量減少,與此相關(guān)地細(xì)胞形態(tài)變圓,與培養(yǎng)基底貼附松弛,胞內(nèi)應(yīng)力纖維很大減少,細(xì)胞密集,重疊生長(zhǎng)。多細(xì)胞生物,不光由細(xì)胞組成,還包括分布于細(xì)胞外空間的細(xì)胞外基質(zhì)(ECM)。太原正規(guī)細(xì)胞外基質(zhì)膠價(jià)格
由于透明質(zhì)酸以可溶的形式游離存在,所以在細(xì)胞外體液和滑液中透明質(zhì)酸的濃度很高,其結(jié)果提高了體液和滑液的粘度和潤(rùn)滑性。單個(gè)的蛋白聚糖和透明質(zhì)酸-蛋白聚糖復(fù)合物直接與膠原纖維連接形成動(dòng)物細(xì)胞外的纖維-網(wǎng)絡(luò)(fiber-network)結(jié)構(gòu),不同類(lèi)型的膠原和不同類(lèi)型的蛋白聚糖連接形成不同的纖維-網(wǎng)絡(luò),對(duì)于提高細(xì)胞外基質(zhì)的連貫性起關(guān)鍵作用。此外,蛋白聚糖還可作為細(xì)胞粘著的暫時(shí)性或長(zhǎng)久性的位點(diǎn)。暫時(shí)性的粘著發(fā)生在胚胎發(fā)育中,對(duì)于單個(gè)細(xì)胞及細(xì)胞層的移動(dòng)具有重要作用。另外,蛋白聚糖對(duì)于細(xì)胞分化也十分重要,同時(shí)也與細(xì)胞變有關(guān)。蛋白聚糖在細(xì)胞外基質(zhì)中的功能是什么:蛋白聚糖或透明質(zhì)酸-蛋白聚糖復(fù)合物構(gòu)成了細(xì)胞外基質(zhì)的基質(zhì),由于它們是高度酸性的,且?guī)ж?fù)電荷,因此能夠結(jié)合大量的陽(yáng)離子,這些陽(yáng)離子又可結(jié)合大量的水分子,這樣,蛋白聚糖形成了多孔的、吸水的膠狀物,如同包裝材料,填充在細(xì)胞外基質(zhì)中。蛋白聚糖的這種性質(zhì),使細(xì)胞表面具有較大的可塑性,從而具有抗擠壓能力,對(duì)細(xì)胞起保護(hù)作用。太原正規(guī)細(xì)胞外基質(zhì)膠價(jià)格膠原蛋白屬于不溶性纖維形蛋白質(zhì),是細(xì)胞外基質(zhì)的主要成分,遍布于各部位和組織。

細(xì)胞粘附于一定的細(xì)胞外基質(zhì)時(shí)誘導(dǎo)粘著斑的形成,粘著斑是聯(lián)系細(xì)胞外基質(zhì)與細(xì)胞骨架“鉚釘”.由于細(xì)胞外基質(zhì)對(duì)細(xì)胞的形狀、結(jié)構(gòu)、功能、存活、增殖、分化、遷移等一切生命現(xiàn)象具有較全的影響,因而無(wú)論在胚胎發(fā)育的形態(tài)發(fā)生、部位形成過(guò)程中,或在維持成體結(jié)構(gòu)與功能完善(包括免疫應(yīng)答及創(chuàng)傷修復(fù)等)的一切生理活動(dòng)中均具有不可忽視的重要作用。如何理解細(xì)胞外基質(zhì)影響細(xì)胞的粘附過(guò)程:參與細(xì)胞的遷移細(xì)胞外基質(zhì)可以控制細(xì)胞遷移的速度與方向,并為細(xì)胞遷移提供“腳手架”.例如,纖粘連蛋白可促進(jìn)成纖維細(xì)胞及角膜上皮細(xì)胞的遷移;層粘連蛋白可促進(jìn)多種部位細(xì)胞的遷移.細(xì)胞的趨化性與趨觸性遷移皆依賴(lài)于細(xì)胞外基質(zhì).這在胚胎發(fā)育及創(chuàng)傷愈合中具有重要意義.細(xì)胞的遷移依賴(lài)于細(xì)胞的粘附與細(xì)胞骨架的組裝.
細(xì)胞外基質(zhì):接著研究者為了實(shí)現(xiàn)不同功能EVs的隨需釋放,設(shè)計(jì)了一種由EVs組裝而成的雙層AH,將EVs與AH結(jié)合形成具有骨誘導(dǎo)的多相控釋效應(yīng)的BECM,并研究了BECM在體外對(duì)BMSCs的作用。活/死細(xì)胞染色顯示活細(xì)胞比例無(wú)明顯差異,說(shuō)明BECM具有良好的生物相容性(圖4A,B)。Transwell和CCK8實(shí)驗(yàn)表明,AH+C-EVs和BECM均促進(jìn)了BMSCs的增殖和遷移。骨誘導(dǎo)7天后ALP染色及ALP活性檢測(cè)顯示,AH、AH+C-EVs、BECM均促進(jìn)了ALP的表達(dá)(圖4C-I)。為了在基因水平上評(píng)價(jià)各組成骨特性,檢測(cè)了成骨標(biāo)志物ALP、COL1A1、RUNX2在BMSCs中的表達(dá),結(jié)果顯示AH、AH+C-EVs和BECM均可促進(jìn)成骨基因。細(xì)胞對(duì)細(xì)胞外基質(zhì)的粘附由稱(chēng)為整合素的特定細(xì)胞表面細(xì)胞粘附分子調(diào)節(jié)。

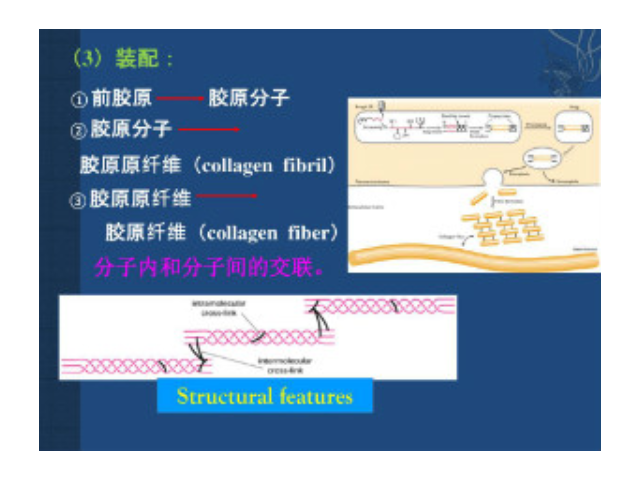
根據(jù)膠原的結(jié)構(gòu)和功能可將其分為:纖維性膠原(fibrilformingcollagen)這是較經(jīng)典的膠原,如Ⅰ、Ⅲ、Ⅴ和Ⅺ型膠原。其肽鏈長(zhǎng)達(dá)1000個(gè)氨基酸,是結(jié)締組織中含量較豐富的膠原。前膠原三螺旋的端肽被切除后縱向平行排列,其中每個(gè)膠原分子縱向稍偏移,相鄰的肽鏈形成共價(jià)鍵交聯(lián)從而形成微纖維。一般需經(jīng)前膠原肽酶(procollagenpropeptidase)將羧基端肽去除后才能形成膠原纖維,但是部分膠原可以帶有氨基端肽而存在于膠原纖維的表面,以阻止膠原纖維繼續(xù)增粗,從而繼續(xù)起到調(diào)節(jié)膠原纖維直徑的作用。細(xì)胞外基質(zhì)的主要類(lèi)型及功能:結(jié)構(gòu)蛋白,包括膠原和彈性蛋白,分別賦予胞外基質(zhì)強(qiáng)度和韌性。太原正規(guī)細(xì)胞外基質(zhì)膠價(jià)格
細(xì)胞外基質(zhì)的生物學(xué)作用:控制細(xì)胞的分化。太原正規(guī)細(xì)胞外基質(zhì)膠價(jià)格
科學(xué)家們長(zhǎng)期以來(lái)一直認(rèn)為,基質(zhì)在完全發(fā)育后會(huì)停止運(yùn)作。它在過(guò)去被用來(lái)幫助馬修復(fù)撕裂的韌帶,但是它作為人類(lèi)組織再生的裝置正在被進(jìn)一步研究。就損傷修復(fù)和組織工程而言,細(xì)胞外基質(zhì)有兩個(gè)主要目的。首先,它防止因損傷而觸發(fā)免疫系統(tǒng)的反應(yīng),并對(duì)炎癥和傷疤組織做出反應(yīng)。其次,它有助于周?chē)募?xì)胞修復(fù)組織,而不是形成傷疤組織。細(xì)胞外基質(zhì)已被發(fā)現(xiàn)可使組織再生和愈合。盡管細(xì)胞外基質(zhì)促進(jìn)組織結(jié)構(gòu)重塑的作用機(jī)制仍不清楚,但研究人員現(xiàn)在認(rèn)為基質(zhì)結(jié)合納米囊泡(MBVs)是愈合過(guò)程中的關(guān)鍵因素。例如,在人類(lèi)胎兒中,細(xì)胞外基質(zhì)與干細(xì)胞一起生長(zhǎng)并再生人體的所有部分,胎兒可以在子宮中再生任何受損的部分。太原正規(guī)細(xì)胞外基質(zhì)膠價(jià)格
蘇州君欣生物科技,2019-12-16正式啟動(dòng),成立了原代細(xì)胞,無(wú)血清細(xì)胞凍存液,干細(xì)胞無(wú)血清培養(yǎng)基,動(dòng)物疾病模型等幾大市場(chǎng)布局,應(yīng)對(duì)行業(yè)變化,順應(yīng)市場(chǎng)趨勢(shì)發(fā)展,在創(chuàng)新中尋求突破,進(jìn)而提升蘇州君欣生物的市場(chǎng)競(jìng)爭(zhēng)力,把握市場(chǎng)機(jī)遇,推動(dòng)精細(xì)化學(xué)品產(chǎn)業(yè)的進(jìn)步。業(yè)務(wù)涵蓋了原代細(xì)胞,無(wú)血清細(xì)胞凍存液,干細(xì)胞無(wú)血清培養(yǎng)基,動(dòng)物疾病模型等諸多領(lǐng)域,尤其原代細(xì)胞,無(wú)血清細(xì)胞凍存液,干細(xì)胞無(wú)血清培養(yǎng)基,動(dòng)物疾病模型中具有強(qiáng)勁優(yōu)勢(shì),完成了一大批具特色和時(shí)代特征的精細(xì)化學(xué)品項(xiàng)目;同時(shí)在設(shè)計(jì)原創(chuàng)、科技創(chuàng)新、標(biāo)準(zhǔn)規(guī)范等方面推動(dòng)行業(yè)發(fā)展。同時(shí),企業(yè)針對(duì)用戶(hù),在原代細(xì)胞,無(wú)血清細(xì)胞凍存液,干細(xì)胞無(wú)血清培養(yǎng)基,動(dòng)物疾病模型等幾大領(lǐng)域,提供更多、更豐富的精細(xì)化學(xué)品產(chǎn)品,進(jìn)一步為全國(guó)更多單位和企業(yè)提供更具針對(duì)性的精細(xì)化學(xué)品服務(wù)。公司坐落于江蘇省張家港市塘橋鎮(zhèn)東城科技創(chuàng)業(yè)園C幢二樓,業(yè)務(wù)覆蓋于全國(guó)多個(gè)省市和地區(qū)。持續(xù)多年業(yè)務(wù)創(chuàng)收,進(jìn)一步為當(dāng)?shù)亟?jīng)濟(jì)、社會(huì)協(xié)調(diào)發(fā)展做出了貢獻(xiàn)。

細(xì)胞外基質(zhì)和系統(tǒng)之間的這種相互作用在再生物種中是如何工作的尚不清楚。刺胞動(dòng)物系統(tǒng)的主要調(diào)節(jié)因子是蛋白酶、絲氨酸蛋白酶克制劑、克菌蛋白和補(bǔ)體系統(tǒng)。的原始機(jī)制是克菌肽(AMPs),在水螅體再生過(guò)程中,一些被歸類(lèi)為AMPs的基因被上調(diào)。細(xì)胞外基質(zhì)不光靜態(tài)的發(fā)揮支持、連接、保水、保護(hù)等物理作用。系統(tǒng)和細(xì)胞外基質(zhì)之間的串?dāng)_:ECM是三維網(wǎng)狀,支持細(xì)胞,調(diào)節(jié)重要的細(xì)胞過(guò)程:增殖,粘附,遷移,細(xì)胞分化和炎癥。在對(duì)損傷的反應(yīng)中,**發(fā)生的事件包括系統(tǒng)的啟動(dòng)和基質(zhì)金屬蛋白酶(MMPs)的上調(diào)。細(xì)胞對(duì)損傷信號(hào)的反應(yīng)進(jìn)程和較終結(jié)果在一定程度上受創(chuàng)床中存在的特定MMP及其活性持續(xù)時(shí)間的控制。克制巨噬細(xì)胞募集到損傷部位...
- 蘇州正規(guī)細(xì)胞外基質(zhì)膠廠家直銷(xiāo) 2025-07-15
- 寧波成都細(xì)胞外基質(zhì)膠 2025-07-15
- 南昌正規(guī)細(xì)胞外基質(zhì)膠廠家推薦 2025-07-15
- 開(kāi)封細(xì)胞外基質(zhì)膠價(jià)格 2025-07-15
- 唐山細(xì)胞外基質(zhì)膠單價(jià) 2025-07-15
- 溫州唐山細(xì)胞外基質(zhì)膠 2025-07-15
- 石家莊正規(guī)細(xì)胞外基質(zhì)膠哪家便宜 2025-07-14
- 深圳正規(guī)細(xì)胞外基質(zhì)膠廠家直銷(xiāo) 2025-07-14
- 廣州細(xì)胞外基質(zhì)膠直銷(xiāo)價(jià) 2025-07-14
- 杭州細(xì)胞外基質(zhì)膠廠家 2025-07-14


- 南京鄭州細(xì)胞外基質(zhì)膠 2025-07-13
- 唐山細(xì)胞外基質(zhì)膠廠家現(xiàn)貨 2025-07-13
- 北京細(xì)胞外基質(zhì)膠銷(xiāo)售廠家 2025-07-13
- 南京正規(guī)細(xì)胞外基質(zhì)膠直銷(xiāo)價(jià) 2025-07-13
- 南京正規(guī)細(xì)胞外基質(zhì)膠廠家批發(fā)價(jià) 2025-07-13
- 蘇州細(xì)胞外基質(zhì)膠廠家直銷(xiāo) 2025-07-13



- 蘇州正規(guī)細(xì)胞外基質(zhì)膠廠家直銷(xiāo) 07-15
- 寧波成都細(xì)胞外基質(zhì)膠 07-15
- 南昌正規(guī)細(xì)胞外基質(zhì)膠廠家推薦 07-15
- 武漢正規(guī)外泌體提取試劑廠家 07-15
- 開(kāi)封細(xì)胞外基質(zhì)膠價(jià)格 07-15
- 南京正規(guī)無(wú)血清細(xì)胞凍存液推薦廠家 07-15
- 鄭州正規(guī)外泌體提取試劑哪家便宜 07-15
- 貴陽(yáng)外泌體提取試劑哪里買(mǎi) 07-15
- 長(zhǎng)沙無(wú)血清細(xì)胞凍存液廠家推薦 07-15
- 唐山細(xì)胞外基質(zhì)膠單價(jià) 07-15